

Authors: Lisa M. Aschemeier, Brian Keane, Nancy G. Solomon
Institution: Miami University of Ohio
Date: February 2008
ABSTRACT
Female mate choice in rodents can be based on genotypic variation among males. Prairie voles (Microtus ochrogaster) exhibit intraspecific variation in the regulatory region of a single gene, avpr1a which encodes the arginine vasopressin 1a receptor (V1aR). Polymorphism in the length of the microsatellite region of the avpr1a gene appears to result in variation in socio-behavioral traits within this species. Males with two long microsatellite alleles have been observed to contribute more time to paternal care and also show a preference toward their partner rather than a strange female relative to males with two short alleles. In this study a three chambered testing apparatus was used. One male with short alleles was tethered in one of the side chambers, and one male with long alleles was tethered in the other side chamber. Female prairie voles were given a choice between the males over a 72 hour period. Choice was based on the total time the female spent with each male and the number of matings with each male. The data is not statistically significant to conclude that females are showing preference toward males with long alleles versus males with short alleles. There is significant data to conclude that females prefer larger males. This data suggests several possibilities that include: females simply cannot differentiate between the differences in the allele lengths, the allele lengths are not the most important factor driving female mate choice, or there is insufficient power at this point to show female preference. Future studies should look at female mate choice in a natural setting and in the lab with body mass of males closely matched. Also, the preference of females from other populations should be examined.
INTRODUCTION
In most vertebrate species, the choice of a mate is thought to be exercised primarily by the female (Majerus 1986). The attributes of males that females prefer vary greatly from species to species and include mating status, dominance status, genetic compatibility, and body size (Solomon and Keane 2007). Females use these traits as indicators of potential direct and indirect benefits such as access to territory and food, reproductive success, or greater fitness of offspring (Solomon and Keane 2007). Females may base mate choice on the material benefits offered by the male. These include the ability of the male to provide her with food and the ability of the male to provide paternal care for her offspring. Mating status of the male might be another important trait in female mate choice. Males that have mated previously in quick succession tend to have a reduced sperm count in later ejaculations (Huck and Lisk 1985; Huck et al. 1986; Austin and Dewsbury 1986). Because of this reduction of fertility in the male, if a female mates with this male, she may risk not being impregnated or give birth to smaller litters; consequently, her overall fitness is reduced.
Females of monogamous species may exhibit greater choosiness than polygamous or promiscuous females (Salo and Dewsbury 1995) because monogamous females usually depend on the male for paternal care; therefore, mating with an unmated male will most likely ensure that the female and her offspring receive all of the male's paternal effort. Studies have shown that in prairie voles females display a social and mating preference for unmated versus mated males (Pierce & Dewsbury 1991). However, field studies have concluded that there is no significant evidence to conclude that female prairie voles discriminate between sexually experienced and sexually inexperienced males (Getz et al. 2004). Also, mating with an unmated male decreases the likeliness of the male to transmit diseases to the female or her offspring (Jennions & Petrie 2000). In prairie voles lab studies have shown that the continued presence of a mate is important for successful reproduction (McGuire et al. 1992; Dewsbury 1995). Dewsbury (1995) also showed that removal of the male partner significantly decreased the likelihood that the female would give birth compared to females whose mates were present.
Females may discriminate between males based on ownership of territory which would provide the female with a safe and secure nest site (Engel 1990). Dominant males offering a high-quality territory may also pass on their dominance status to their offspring (Agrell et al. 1998; Drickamer 1992). Females may also be able to discriminate between males with different genotypes, therefore choosing males with genes that will be beneficial to her or her offspring (Drickamer et al. 2000; Lenington 1983). Several studies have provided evidence that females can discriminate between potential mates based on polymorphism at a single locus. In inbred strains of laboratory mice, females sometimes use single gene differences in MHC haplotypes as a basis for mate choice to apparently avoid further inbreeding (Egid & Brown 1989; Arcaro & Elkund 1998). In house mice it has been observed that females heterozygous at the t-locus (Tt) preferentially mate with homozygous wild-type males (TT) and thus avoid producing inviable t/t offspring (Lenington 1991; Lenington et al. 1994). It has been shown in a study by Kavaliers et al. (2004) that female mice were able to discriminate between odors of male mice with the gene for oxytocin or estrogen receptor α or β knocked out as compared to their wild-type littermates.
This study examines the effect the length of the microsatellite of the avpr1a gene has on female mate choice in prairie voles. In voles, studies have shown that V1aR expression is regulated by the microsatellite regulatory region of avpr1a (Young 1999; Hammock & Young 2005). Vasopressin is a neuropeptide which regulates the expression of some social behaviors (Young 1999; Goodson and Bass 2001; Phelps and Young 2003). In vertebrates, vasopressin is associated with reproductive behaviors, such as paternal care, affiliative behavior, and pair-bond formation (Young and Wang 2004; Wang and Aragona 2004; Hammock et al. 2005). Differences in the expression of the V1aR underlie intraspecific variation in male social behavior. In prairie voles, a monogamous rodent, males show intrapopulation variation in the length of the microsatellite region within the gene encoding V1aR, and these polymorphisms are correlated with differences in behavior (Hammock and Young 2005). Longer avpr1a microsatellites cause an increased amount of V1aR expression. Hammock and Young (2005) observed, in laboratory tests, that male prairie voles with longer avpr1a microsatellite alleles displayed a significant preference for their partner compared to an unfamiliar female. Therefore, variations in avpr1a microsatellite length appear to affect the degree of social attachment of male prairie voles to their mates (Young et al. 1997; Hammock and Young 2005). Also, males with longer avpr1a microsatellite alleles displayed significantly higher levels of parental care.
The purpose of this study is to determine if female prairie voles discriminate between males based on the length of a male's alleles at the avpr1a microsatellite. Based on the results of Hammock and Young's (2005) study, we predicted that if females can discriminate between males based on the length at the avpr1a microsatellite, they should prefer males with longer alleles relative to males with short alleles since males with long alleles should be more faithful and provide more paternal care to offspring. By examining the potential effects that polymorphism in the V1aR gene has on female mate choice, one aspect of sexual selection, this study will generate some mechanistic and evolutionary insights into the intraspecific natural variation in a complex social behavior such as monogamy.
MATERIALS AND METHODS
Animals
All prairie voles used in this experiment were F2 generation animals produced from wild adult prairie voles originally live-trapped in east-central Illinois. The initial body mass and age of each male vole was obtained before the start of this experiment. They were housed in the wild animal facility at Miami University in Oxford, OH. Voles were housed in polycarbonate cages (28 x 22 x 15 cm or 38 x 33 x 17 cm). Recycled paper bedding (Carefresh; Fangman Industries, Cincinnati, OH), alfalfa, and cotton Nestlets (Ancare Inc.) were used as nesting material. Voles were fed ad lib food (PIM Nutrition International's 5013 Rodent Breeder Diet, Brentwood, MO) and water. Voles were also fed rabbit chow (Lab Diet's 5326 High Fiber Rabbit) once weekly. Temperature was maintained at 20±1oC. Room lighting was provided by over head fluorescent lights on a 14:10h photoperiod with lights on at 6 h.
Using tissue from toes and ear, males were genotyped to determine the lengths of their avpr1a microsatellite alleles. To determine the length of the avpr1a microsatellite, PCR was used to amplify the region of the V1aR gene with primers specific to prairie voles (Hammock and Young 2005). Only males were genotyped at this locus. To verify that the correct region of avpr1a was being amplified, the microsatellite was sequenced.
PCRs for the avpr1a locus were 15 µl reactions with 100 ng/µl DNA, 10 mM Tris-HCL, 0.2 mM dNTPs, 1.5 mM MgCl, 0.67 µM of both forward (fluorescently labeled with 6-FAM phosphoramidite (IDT DNA Technologies, Coralville, IA)) and reverse primers, and 0.5 U Taq 11 DNA polymerase in its supplied reaction buffer (Promega, Madison, WI). Initial denaturation took place for 2 minutes at 94oC, subsequent PCR amplification took place in a Biometra thermal cycler for 25 cycles: 10x (94oC, 15 s; 57oC, 30 s; 72oC, 45 s); 15x (94oC, 15 s; 57oC, 30 s; 72oC, 45 s) with final elongation for 7 minutes at 72oC.
The amplified products of the PCRs were diluted and combined with an internal size standard (ROX GS2500, Applied Biosystems, Foster City, CA) and denatured. Fragment length (molecular weight) measurements were conducted on an ABI 3730 sequencer run for 38 minutes (dye set D) with a 50 cm capillary (Applied Biosystems, Foster City, CA).
Fragment length analysis was conducted using GeneMapper software (Applied
Biosystems, Foster City, CA). Fragment lengths were compiled into the same 5 base pair bins as described by Hammock and Young (2005). If both alleles were less than or equal to the median allele length bin (733), these males were classified as having "short", and males with allele lengths greater than the median allele length bin were classified as those with "long" alleles.
Fourteen sexually naïve females were tested in this experiment. A total of eight males were used as stimulus animals, four with two short alleles (719.2-723.7 bp) and four with two long alleles (750-771 bp).
Each male was used in at least two and no more than five trials. Males were matched to form a stimulus set based on age and the length of avpr1a microsatellite. Body mass of males was also considered when forming stimulus sets, but, due to limited number of males some pairs had males that differed by up to 10g.
Apparatus
The experiment was conducted in a three-chambered apparatus (20.5 x 60cm). This apparatus consisted of a Plexiglas box divided into three equally-sized chambers. Each of the chambers was (20 x 20.5cm). Plexiglas dividers were used to separate the three compartments. The dividers had a hole (3.5cm diameter) in the bottom which allowed the female to move freely among all compartments. Food, water, a Nestlet, and recycled paper bedding were placed in each chamber and were checked every day during each trial. Water, food, and Nestlet were replaced, if necessary, in chambers that did not have any during that day. The photoperiod and room temperature in the room where the testing was done were maintained as in the lab colony. The apparatus was cleaned using a mixture of water and Quatricide mixture after the completion of each trial.
Test Trials
Before each trial, two males, one with two short alleles and one with two long alleles for the avpr1a microsatellite, were selected for each experiment. The males were collared using a plastic cable-tie (Diversified Supply, Inc.; Cincinnati, OH). The cable-tie was then attached to a steel fishing leader (WalMart) which was then attached to the wire mesh at the top of the cage. Each male was placed in one of the end chambers. The males had free movement within their chambers but could not enter into the middle chamber or directly interact with each other. Males were collared and placed in the choice arena approximately 12 hours before the start of the trial. Position of male with two long alleles was alternated in side chambers during every trial to account for the possibility of a position effect.

Figure 1. Time spent by females with male prairie voles based on length of avpr1a alleles. The central line within each box is the median, and the bottom and top of each box represent the 25th and 75th percentiles, respectively. Whiskers denote the minimum and maximum values.
Following the approximate 12 hour acclimation period, a sexually naïve female, that was unrelated to the two males, was placed in the center chamber. The events that followed were videotaped for the next 72 hours using a time-lapse VCR (Panasonic AG6040) with one frame per six seconds. Two lamps with 25 W red light bulbs were used during the times when the lights were off in the building. After recording was completed, the voles were returned to their home cages.
Each video was watched in its entirety. The amount of time the female spent in each male's chamber and the number of copulations were recorded along with movement of food and bedding by the female and possible copulations. A social preference was defined as the male with which the female spent the most time. Mating preference was defined as the male that the female copulated with most frequently.
Data Analysis
A Wilcoxon Signed Rank Test was used to analyze female social preference based on time spent with males with long alleles versus males with short alleles and mating preference based on number the of matings with males with long alleles versus males with short alleles. Because male body mass was not controlled for the pairings used in the experiment, a Wilcoxon Signed Rank Test was used to determine if male body mass influence the time spent with each male and, also, number of times mated with each male. All values are reported as means ± SE. Results were considered statistically significant if p less than or equal to 0.05.
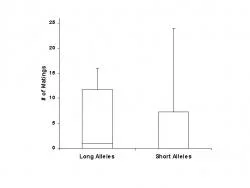
Figure 2. Number of matings by females with male prairie voles based on lengths of the avpr1a alleles. The central line within each box is the median, and the bottom and top of each box represent the 25th and 75th percentiles, respectively. Whiskers denote the minimum and maximum values.

Figure 3. Body mass of preferred and non-preferred male prairie voles based on time spent by females. The central line within each box is the median, and the bottom and top of each box represent the 25th and 75th percentiles, respectively. Whiskers denote the minimum and maximum values.
RESULTS
In all 14 trials, females visited both males during the 72 hour trial period. Females did spend a majority of their time in either of the males' chambers compared to the time spent in the middle chamber. Females did not show a social preference for males based on their avpr1a microsatellite allele lengths (p = 0.477, n = 14, Figure 1). Mean time females spent with males with short alleles was 1881.43±338.72 minutes and mean time females spent with males with long alleles was 1917.93±332.02 minutes. Of the eleven females that mated, most females only mated with one of the males (9 of 11, 81.8%); however, some females mated with both males (2 of 11, 18.2%). Three females mated with neither male (3 of 14, 21.4%). Mean number of matings of females with males with short alleles was 5.07±2.31 and 5.21±1.79 matings by females with males having long alleles. Females did not show a mating preference for males based on avpr1a microsatellite allele lengths (p = 0.859, n = 14, Figure 2). Females did show a social and mating preference for the male with larger body mass (Social Preference: Figure 3; Mating Preference: Figure 4). Females showed a significant social preference for the male with larger body masses (p = 0.032, n = 14), and a statistically significant mating preference for males with larger body masses (p = 0.035, n =14). For socially preferred males the mean weight was 58.04±2.65g compared to the non-preferred males with a mean weight of 50.60±2.46g. For males that females preferred to mate, the mean weight was 57.48±2.09g compared to the non-preferred males with a mean weight of 52.37±2.41g. Reported standard errors for this study are very large; however, even in the simplest situations, behavior is complex and highly variable (Ploger & Yasukawa 2003).

Figure 4. Body mass of preferred and non-preferred male prairie voles based on number of matings with males by females. The central line within each box is the median, and the bottom and top of each box represent the 25th and 75th percentiles, respectively. Whiskers denote the minimum and maximum values.
DISCUSSION
We had expected that avpr1a microsatellite length in male prairie voles would be important for females since males with long alleles should be more faithful and provide more paternal care to offspring (Hammock and Young 2005). However, no evidence was found to support the prediction that females prefer males with longer alleles as social or mating partners. These results suggest that female prairie voles do not show preference based on avpr1a microsatellite allele length. There are several different possibilities of why females show no preference which include the following: females simply cannot differentiate between the differences in the microsatellite allele lengths, the microsatellite allele lengths are not the most important factor driving female mate choice, or there is insufficient power to show female preference due to small sample size.
Females in our study may have not been able to distinguish the difference between males based on the allele lengths. Hammock and Young (2005) found that males with different allele lengths show differences in behavior. Perhaps, because males were tethered in our study, they were unable to show their potential faithfulness to the female. Therefore, the female was unable to discriminate between allele lengths.
Female mate choice can be based on several different characteristics of the male which include mating status, dominance status, genetic compatibility, body size, or spatial ability (Solomon and Keane 2007). Perhaps, in our study, females considered other characteristics to be more important than the allele lengths. Females may look at several factors when choosing a mate with some characteristics carrying more importance than others (Spritzer et al. 2005).
In our study, females did show a preference for males with larger body mass. This is consistent with previous findings that, in the lab, both prairie vole males and females preferred to associate with opposite-sex conspecifics that were heavier at weaning (Solomon 1993). However, in the field there was no evidence found to suggest that female prairie voles based their choice of mate on body mass or condition (Getz et al. 2004, Keane et al. 2007). This difference between lab and field results may be caused by the differences in the experimental settings.
There is another possible explanation for the behavior that we observed in this experiment. Fink et al. (2006) recently suggested that mammalian monogamy is not controlled by avpr1a alone. They found the avpr1a microsatellite in 19 of 21 Microtus species in which only three species exhibit social monogamy (Microtus ochrogaster, Microtus pinetorum, and Microtus multiplex). In response to this study, Young and Hammock (2007) suggest that polymorphisms in the avpr1a microsatellite tweak the brain avpr1a expression patterns and in turn social behavior tendencies. Insel et al. (1994) found that the distribution of V1aR in the brain differs between the socially monogamous and socially non-monogamous species. Young et al (1999) found that prairie vole and pine vole avpr1a loci contain 430-base complex microsatellite sequence, which is composed of simple sequence repeats interspersed with nonrepetitive elements. They found that this microsatellite is found to be expressed in a truncated version in the avpr1a loci of the nonmonogamous montane and meadow voles. As found by Hammock and Young (2004) an increase of V1aR in the brain is caused by the expansion of avpr1a microsatellite. Hammock and Young (2004) suggest that this difference between monogamous and nonmonogamous species may account for the species differences in brain expression of avpr1a and mating strategy.
Our results showed a lack of association between female mate preference and male avpr1a microsatellite allele lengths and caused us to reject of our hypothesis. Future studies that may support these findings might include studies with larger sample size. In our study, females spent more time with males with long alleles but the sample size used in this study may not have been large enough to detect a statistically significant difference. Also, body mass needs to be matched more closely since we found that females use this trait in social and mating preferences. Due to the limited number of males available for this study, males were matched as closely as possible for body mass. A greater difference in male genotypes could be used in future studies. Also, observations from more realistic studies such as one in a natural setting could be a prospect for further research.
Our study looked at female mate choice based on the male's avpr1a micrcosatellite allele length, but we found that females chose males based on body size. In order to discover if females choose males based on avpr1a microsatellite allele length, future studies should control for male body size. However, our findings with body size could also be a basis for future research in prairie voles and other species. Based on conflicting evidence from lab studies and field studies (Solomon 1993; Getz et al. 2004; Keane et al. 2007), the impact that body size and/or condition has on female mate choice should be further studied. Male body size could display a male's physical condition which is strongly correlated with good genes (Solomon & Keane 2007).
Overall, the results from this experiment demonstrate that avpr1a microsatellite allele length has no significant effect on female mate choice in a laboratory setting for prairie voles when members of the stimulus set differ in body mass. The next step to further test this experimental result is to analyze female mate choice in a natural setting and in the lab with body mass of males closely matched and, also, to examine preferences of female prairie voles from other populations. This research allows us to better understand female mate choice and the effects that differing lengths in avpr1a microsatellite alleles have on that choice.
ACKNOWLEDGEMENTS
We would like to thank Ashley Richmond, Kristen Lucia, and Frank Castelli for their assistance on this research project. Nancy Solomon and Brian Keane were supported by NSF DEB-0316818, and Lisa Aschemeier was supported by REU-NSF DBI-0353915.
REFERENCES
Agrell, J. et al. (1998) Counter-strategies to infancticide in mammals: Costs and consequences. Oikos 83, 507-517.
Arcaro, K.F. and A. Eklund. (1998) A review of MHC-based mate preferences and fostering experiments in two congenic strains of mice. Genetica 104, 241-244.
Austin, D. and D.A. Dewsbury. (1986) Reproductive capacity of male laboratory rats. Physiology and Behavior 37, 627-632.
Dewsbury, D.A. (1995) Role of male proximity in pregnancy maintenance in prairie voles, Microtus ochrogaster. Physiology & Behavior 57, 827-829.
Drickamer, L.C. (1992) Oestrous female house mice discriminate dominant from subordinate males and sons of dominant from sons of subordinate males by odor cues. Animal Behaviour 43, 868-870.
Drickamer, L.C. et al. (2000) Free female mate choice in house mice affects reproductive success and offspring viability and performance. Animal Behaviour 59, 371-378.
Egid, K. and J.L. Brown. (1989) The major histocompatability complex and female mating preferences in mice. Animal Behaviour 38, 548-550.
Engel, C.R. (1990) Scent Marking, Residency, and Female Choice in the European Rabbit (Oryctolagus cuniculus). In: Chemical Signals in Vertebrates. Vol. 5. Ed. By D.W. Macdonald et al. pp. 329-335. Oxford: Oxford University Press.
Fink, S. et al. (2006) Mammalian monogamy is not controlled by a single gene. Proceedings of the National Academy of Sciences 103, 10956-10960.
Getz, L.L. et al. (2004) Does mate choice take place in free-living prairie voles Microtus ochrogaster? Evidence from field data. Acta Zoologica Sinica 50(4), 527-534.
Goodson, J.L. and A.H. Bass. (2001) Social behavior functions and related anatomical characteristics of vasotocin/vasopressin systems in vertebrates. Brain Research Reviews 35, 246-265.
Hammock, E.A.D. and L.J. Young (2004) Functional microsatellite polymorphisms associated with divergent social structure in vole species. Molecular Biology and Evolution 21, 1057-1063.
Hammock, E.A.D. and L.J. Young. (2005) Microsatellite instability generates diversity in brain and sociobehavioral traits. Science 308, 1630-1634.
Hammock, E.A.D. et al. (2005) Association of vasopressin 1a receptor levels with a regulatory microsatellite and behavior. Genes, Brain and Behavior 4, 289-301.
Huck, U.W. and R.D. Lisk. (1985) Determinants of mating success in the golden hamster (Mesocricetus auratus): I. Male capacity. Journal of Comparative Psychology 99, 98-107.
Huck, U.W. et al. (1986) Determinants of mating success in the golden hamster (Mesocricetus auratus): Social dominance and mating tactics under seminatural conditions. Animal Behaviour 34, 971-989.
Insel, T.R. et al. (1994) Patterns of brain vasopressin receptor distribution associated with social organization in microtine rodents. Journal of Neuroscience 14, 5381-5392.
Jennions, M.D. and M. Petrie. (2000) Why do females mate multiply? A review of the genetic benefits. Biological Review 75,21-64.
Kavaliers, M. et al. (2004) Oxytocin and estrogen receptor alpha and beta knockout mice provide discriminably different odor cues in behavioral assays. Genes Brain and Behavior 3, 189-195.
Keane, B. et al. (2007) Effect of body condition at weaning on survival and reproduction in prairie voles. Canadian Journal of Zoology 85, 718-727.
Lenington, S. (1983) Social preferences for partners carrying good genes' in wild house mice. Animal Behaviour 31, 325-333.
Lenington, S. (1991) The t complex: a story of genes, behavior, and populations. Advances in the Study of Behavior 20, 51-86.
Lenington, S. et al. (1994) Female preference and variability among t-haplotypes in wild house mice. The American Naturalist 143, 766-784.
Majerus, M.E.N. (1986) The genetics and evolution of female choice. Trends in Ecology and Evolution 1, 1-7.
McGuire, B. et al. (1992) The effects of mate removal on pregnancy success in prairie vole (Microtus ochrogaster) and meadow voles (Microtus pennsylvanicus). Biology of Reproduction 47,37-42.
Phelps, S.M. and L.J. Young. (2003) Extraordinary diversity in vasopressin (V1a) receptor distributions among wild prairie voles (Microtus ochrogaster): patterns of variation and covariation. Journal of Comparative Neurology 466, 564-576.
Pierce, J.D., Jr. and D.A. Dewsbury. (1991) Female preferences for unmated versus mated males in two species of voles (Microtus ochrogaster and Microtus montanus). Journal of Comparative Psychology 105, 165-171.
Ploger, B.J. and K. Yasukawa. (2003) Introduction to Statistics. In: Exploring Animal Behavior in Laboratory and Field: A Hypothesis-Testing Approach to the Development, Causation, Function, and Evolution of Animal Behavior. Ed. By B.J. Plogger and K. Yasukawa. San Diego: Academic Press.
Salo, A.L. and D.A. Dewsbury. (1995) Three experiments on mate choice in meadow voles (Microtus pennsylvanicus). Journal of Comparative Psychology 109, 42-46.
Solomon, N.G. (1993) Body size and social preferences of male and female prairie voles, Microtus ochrogaster. Animal Behaviour 45, 1031-1033.
Solomon, N.G. and B. Keane. (2007) Reproductive Strategies in Female Rodents. In: Rodent Societies: An Ecological and Evolutionary Perspective. Ed. By J.O. Wolff and P. Shermann. pp. 42-56. Chicago: University of Chicago Press.
Spritzer, M.D. et al. (2005) Female choice based on male spatial ability and aggressiveness among meadow voles. Animal Behaviour 69, 1121-1130.
Wang, Z and B. Aragona. (2004) Neurochemical regulation of pair bonding in male prairie voles. Physiology & Behavior 83, 319-328.
Young, L.J. (1999) Oxytocin and vasopressin receptors and species typical-social behaviors. Hormones and Behavior 36, 212-221.
Young, L.J. and E.A.D. Hammock. (2007) On switches and knobs, microsatellites and monogamy. Trends in Genetics 23, 209-212.
Young, L.J. and Z. Wang. (2004) The neurobiology of pair bonding. Nature Neuroscience. 7, 1048-1054.
Young, L.J. et al. (1997) Species differences in Va1 receptor gene expression in monogamous and nonmonogamous voles: behavioral consequences. Behavioral Neuroscience 111, 599-605.
Young, L.J. et al. (1999) Increased affiliative response to vasopressin in mice expressing the vasopression receptor from a monogamous vole. Nature 400, 766-768.